Medical Community Information
Technical information for medical professionals covering the following topics:
Background, Clinical Presentation, Diagnosis, Management and Treatment, and Molecular Mechanism of Action.
This information is courtesy of Lora E. Fleming, Director of the European Centre for Environment and Human Health and Chair of Oceans, Epidemiology and Human Health at the University of Exeter Medical School
ASP is a newly identified marine toxin disease, first reported from Canada and later identified as a continuing problem in Washington State and Oregon. After an initial gastroenteritis with neurologic symptoms, some persons with ASP develop apparent permanent neurologic deficits, especially dementia. It is caused by the consumption of contaminated shellfish.
At the end of November 1987, 153 cases of acute intoxication after ingestion of toxic mussels were reported in Canada associated with blooms of the marine diatom Pseudonitschia pungens. Dinoflagellate samples taken from the Cardigan River estuary were found to contain >1% domoic acid dry weight. Contaminated mussels had digestive glands engorged with P. pungens.
Clinical Presentation:
Acute symptoms of ASP included vomiting, diarrhea, and in some cases, there followed confusion, loss of memory, disorientation and even coma. The acute symptoms were mild compared with PSP. Permanent neurologic sequelae, especially cognitive disfunction, were most likely in persons who developed neurologic illness with 48 hours, males, in older patients (> 60 yrs), and in younger persons with pre-existing illnesses such as diabetes, chronic renal disease and hypertension with a history of transient ischemic attacks. Three elderly patients died directly and one died indirectly from the intoxication; 19 people required hospitalization with 12 admitted to intensive care units due to seizures, coma, profuse respiratory secretions or unstable blood pressure.
The acute symptom frequencies were the following: vomiting (76%), abdominal cramps (50%), diarrhea (4%), severe headache (43%), and loss of short-term memory (25%). Acutely, the patients had headache, hyporeflexia, hemiparesis, ophthalmoplegia and abnormalities of arousal ranging from agitation to coma; in addition; seizures and myoclonus were observed acutely, especially around the face.
Teitlebaum et al. (1990) studied 14 persons with severe neurologic disease. In neuropsychological testing performed several months after the acute episode, 12/14 persons had severe antegrade memory deficits with relative preservation of other cognitive functions. 11/14 persons had clinical and electromyographic evidence of pure motor or sensory motor neuronopathy or axonopathy. The PET results in 4/14 persons showed decreased glucose metabolism in the medial temporal lobes. The neuropathology for the 4 fatal cases, revealed neuronal necrosis and loss, predominantly in the hippocampus and amygdala. All 14 with severe neurologic disease reported confusion and disorientation within 1.5 to 48 hours after consumption. The maximal neurologic deficits were seen 4 hours post ingestion of least affected and 72 hours in those most affected, with maximal improvement 24 hours to 12 weeks post ingestion. Acute coma was associated with the slowest recovery. Seizures ceased by 4 months but were frequent upto 8 weeks.
Relative preservation of intellect and higher cortical function appears to distinguish this disease from Alzheimer's Disease, and the absence of confabulation with well preserved frontal lobe function is not typical of Korsakoff's syndrome.
Diagnosis:
Management and Treatment:
At this point, the treatment of ASP is symptomatic and supportive. Teitelbaum et al (1990) noted that the seizures respond to iv diazepam and phenobarbital. Three patients were resistent to dilantin for seizure control.
As with many of the marine toxin induced diseases, the initial or index case(s) are often the tip of the iceberg. Therefore any suspected cases of ASP should be reported to the appropriate public health authorities for follow up to ascertain other cases and to prevent further spread. And every effort should be made to obtain contaminated materials and their source.
Since an estimated concentration of 200 ug/g wet weight domoic acid appeared to affect some consumers, with a safety factor of 0.1 applied, Canada has set a concentration of domoic acid of 20 ug/g wet weight above which shellfish commercial operations should be closed. Finally, this epidemic has lead to new attention to the diatoms, especially the appearance of the mucilage from diatoms with species of Nitzschia.
Molecular Mechanism of Action:
Domoic acid, and its co-existing natural chemical analogs act as a potent excitatory neurotransmitter. An estimated concentration of 200 ug/g wet weight domoic acid appeared to affect some human consumers.
Domoic acid is heat-stable and similar to its biochemical analogues, kainic acid and glutamic acid and binds at the same receptor site in CNS. Lesions in human brain, especially in the hippocampus, have been reported in the ASP human cases which are similar to those seen in rats after kainic acid iv administration. When rats are exposed experimentally to domoic acid and its analogues, they get limbic seizures, memory and gait abnormalities, and degeneration of the hippocampus. In animals, domoic acid is three times more potent than kainic acid and 30-100 more potent than glutamic acid.
Recent work by Novelli et al (1992) demonstrated that domoic acid from mussels is more neurotoxic for cultured human neurons than purified domoic acid. This increase is believed to be due to domoic acid potentiation, even in subtoxic amounts, of the excitotoxic effect of glutamic acid and aspartic acid. Glutamic and aspartic acids are present in high concentrations in mussel tissue. This neurotoxic synergism may occur through a reduction in the voltage dependent Mg2+ block at the NMDA receptor associated channel, following activation of non-NMDA receptors by domoic acid.
In humans, domoic acid appears to cause a non-progressive acute neuronopathy involving anterior horn cells or a diffuse axonopathy predominantly affecting motor axons. The acute neuronal hyperexcitation syndrome presumably results from the stimulus of central and possibly peripheral neurons, followed by chronic loss of function in neural systems susceptible to excitotoxic degeneration (ie. hippocampus and anterior horn cells of spinal cord).
References:
- Baden D., L.E. Fleming, and J.A. Bean. 1995. Chapter: Marine Toxins. in: Handbook of Clinical Neurology: Intoxications of the Nervous System Part II. Natural Toxins and Drugs. FA deWolff (Ed). Amsterdam: Elsevier Press, pgs. 141-175.
- Debonnel, G., L. Beaushesne and C. Demonigny. 1989. Domoic acid, the alleged mussel toxin, might produce ite neurotoxic effect through kainate receptor acitivation: an electrophysiologic study in the rat dorsal hippocampus. Canadian Journal of Physiology and Pharmacology 67: 29-33.
- Lawrence, J.F., C.F. Charbonneau, C. Menard, M.A. Quilliam and P.G. Sim. 1989. Liquid chromatographic determination of domoic acid in shellfish products using the paralytic shellfish poison extraction procedure of the Association of Official Analytical Chemists. Journal of Chromatography 462: 349-356.
- Novelli, A., J. Kispert, T. Fernandez-Sanchez, A. Torreblanca and V. Zitko. 1992. Domoic acid-containing toxic mussels produce neurotoxicity in neuronal cultures through a synergism between excitatory amino acids. Brain Research 577: 41-48.
- Perl, T.M., L. Bedard, T. Kosatsky, J.C. Hockin, E.C.D. Todd and R. Remis. 1990. Encephalopathy Caused by Contaminated Mussels - Perl et al. New England Medical Journal 322: 1775-80.
- Subba Rao, D.V., M.A. Quilliam and R. Pocklington. 1988. Domoic acid - a neurotoxic amino acid produced by the marine diatom Nitzschia pungensin culture. Canadian Journal of Fisheries and Aquatic Science 45: 2076-2079.
- Teitelbaum, J.S., R.J. Zatorre, S. Carpenter, D. Gendron, A.C. Evans, A. Gjedde and N.R. Cashman. 1990. Neurologic sequelae of domoic acid intoxication due to the ingestion of contaminated mussels. New England Journal of Medicine 322: 1781-87.
- Wright, J.L.C., R.K. Boyd, A.S.W. De Freitas et al. 1989. Identification of ic acid, a neuroexcitatory amino acid, in toxic mussels from eastern Prince Edward Island. Canadian Journal of Chemistry 67: 481-490.
This information is courtesy of Lora E. Fleming, Director of the European Centre for Environment and Human Health and Chair of Oceans, Epidemiology and Human Health at the University of Exeter Medical School
The most commonly reported marine toxin disease in the world is Ciguatera, associated with consumption of contaminated reef fish such as barracuda, grouper, and snapper. Under-diagnosis and under-reporting (especially in endemic areas such as the Caribbean) make it difficult to know the true worldwide incidence of the Marine Toxin Diseases. At least 50,000 people per year who live in or visit tropical and subtropical areas suffer from Ciguatera worldwide. For example, CDC and others estimate that only 2-10% of Ciguatera cases are actually reported in the United States. In the US Virgin Islands, there are an estimated 300 cases per 10,000 or 3% of the population per year; a similar rate is found in the French West Indies. In St. Thomas, a household survey estimated that 4.4% of all households suffered from Ciguatera annually (at least 2640 persons per year or an annual incidence of 600 cases per year); in Puerto Rico, 7% of the residents have experienced at least one episode of Ciguatera in their lifetime.
Ciguatera, especially in the Caribbean, suffer for weeks to months with debilitating neurologic symptoms, including profound weakness, temperature sensation changes, pain, and numbness in the extremities. Dinoflagellates in the genus Gambierdiscus produce ciguatoxin precursors, and are distributed throughout tropical and sub-tropical regions of the world.
Clinical Presentation:
Ciguatera presents primarily as an acute neurologic disease manifested by a constellation of gastrointestinal (diarrhea, abdominal cramps and vomiting), neurologic (paresthesias, pain in the teeth, pain on urination, blurred vision, temperature reversal) and cardiovascular (arrhythmias, heart block) signs and symptoms within a few hours of contaminated fish ingestion. The pathneumonic symptom of Ciguatera intoxication is hot/cold temperature reversal, although not all patients report this.
The attack rate has been reported to be 73%-100% with ingestion of contaminated fish, without any apparent age-related susceptibility. Acute fatality, usually due to respiratory failure, circulatory collapse or arrhythmias, ranges from 0.1% to 12% of reported cases; presently in the Pacific, the mortality is less than 1%. Lethality is usually seen with ingestion of the most toxic parts of fish (ie. the liver, viscera, roe and other organs).
The clinical picture may be variable among individuals, even with the same food source, different ethnic groups, and possibly with different types of fish and/or geographic location. It appears that ciguatera from consumption of carnivore species may be more toxic than that from consumption of herbivores due to exposure to more than one toxin and/or transformation of the toxin(s) and/or an increased dose response. In Polynesia, Ciguatera is dominated and initiated by neurologic symptoms (90% of patients report paresthesias and dysesthesia), while reports from the Caribbean suggest that Ciguatera initially presents acutely as a gastroenteritis often with associated cardiovascular symptoms, with the gradual onset and dominance of neurologic symptoms over the first 24 hours. This may be due to different toxins mixtures elaborated by Caribbean and Polynesian Gambierdiscus spp.
The symptoms of Ciguatera poisoning, especially the paresthesias and weakness, can persist in varying severity for weeks to months after the acute illness. Prolonged itching due to chronic Ciguatera can present as a dermatologic disease when it is really due to ciguatera paresthesias. Chronic ciguatera can also present as a psychiatric disorder of general malaise, depression, headaches, muscular aches, and peculiar feelings in extremities for several weeks. It is reported that those with chronic symptoms seem to have recurrences of their symptoms with the ingestion of fish (regardless of type), ethanol, caffeine, and nuts 3 to 6 months from initial ingestion.
Ciguatera can be sexually transmitted. With exposure of the mother, premature labor and spontaneous abortion have been reported, as well as effects on the fetus and newborn child through placental and breast milk transmission.
In the differential diagnosis of Ciguatera, poisoning with the other marine toxins, especially NSP and PSP should be considered since dysesthesias with nausea, vomiting and diarrhea are the presenting symptoms. Obviously the history of fish versus shellfish consumption should help to differentiate. Type E botulism with ingestion of smoked fish, Scromboid and even Eosinophilic meningitis from helminthic infection of Angiostrongylus cantonensis from ingestion of raw mollusks, crabs and certain fish should be considered. Finally, poisoning with organophosphates pesticides can present initially with a similar clinical picture except for the exposure history.
Diagnosis:
Management and Treatment:
Medical treatment has been to a large extent symptomatic; a variety of agents, including vitamins, antihistamines, anticholinesterases, steroids and tricyclic antidepressants, have been tried with limited results. Gut emptying and decontamination with charcoal is recommended acutely although often the severe ongoing vomiting and diarrhea prevents this. Atropine is indicated for bradycardia, and dopamine or calcium gluconate for shock. It is recommend that opiates and barbiturates be avoided since they may cause hypotension, and opiates may interact with maitotoxin.
With apparent considerable success, at least acutely, mannitol infusions have been used. Palafox et al (1988) administered 1 gm/kg of 20% mannitol at a rate of 500 mL/h "piggybacked" to an iv infusion of 5% dextrose in Ringers lactate or saline solution at 30 mL/h or more depending on fluid requirements with complete reversal of symptoms in the majority of patients tested. Subsequent reports have affirmed his success although mannitol appears to be most effective in completely relieving symptoms when given within the first 48-72 hours from ingestion.
Amitriptyline (25 to 75 mg bid) and similar medications do seem to have some success in relieving the symptoms of chronic Ciguatera, such as fatigue and paresthesias. It is possible that nifedipine may be appropriate as a calcium channel blocker to counteract the effects of maitotoxin. Finally, there are over 64 different local remedies including medicinal teas used in both the Indo-Pacific and West Indies regions. None of these treatments have been evaluated in a controlled clinical trial with the exception of two controlled trials of Mannitol for treatment of acute Ciguatera, so that their true efficacy is impossible to determine.
As mentioned above, there appears to be a sensitivity to certain foods (ie. ingestion of fish (regardless of type), ethanol, caffeine, and nuts) after ciguatera poisoning and these should be avoided for 3 to 6 months after the illnesses. In addition, there is no immunity to this illnesses and recurrences of actual ciguatera in the same individual appear to be worse than the initial illness.
As with many of the marine toxin induced diseases, the initial or index case(s) are often the tip of the iceberg. Therefore any suspected cases of Ciguatera should be reported to the appropriate public health authorities for follow up to ascertain other cases and to prevent further spread. And every effort should be made to obtain contaminated materials and their source.
Obviously persons who live in or travel to endemic areas should never eat barracuda or morey eel, and should be cautious with grouper and red snapper, as well as enquiring about local fish associated with Ciguatera. Since there is no reliable way to "decontaminate" or even to distinguish contaminated fish by smell or appearance, at a minimum, people should be advised to avoid the viscera of any reef fish as well as avoiding consuming unusually large predacious reef fish especially during the reproductive season.
Molecular Mechanism of Action:
The two most common toxins associated with Ciguatera are Ciguatoxin and Maitotoxin, and they are some of the most lethal natural substances known. In mice, ciguatoxin is lethal at 0.45 ug/kg ip, and maitotoxin at a dose of 0.15 ug/kg ip. Oral intake of as little as 0.1 ug ciguatoxin can cause illness in the human adult (as an extrapolation from fish samples eaten).
Ciguatoxin, a lipid soluble substance, opens voltage dependant sodium channels in cell membranes which induces membrane depolarization. It causes prolonged symptoms indicate nerve blockage or damage requiring regeneration of nervous tissue Maitotoxin, water soluble, specifically increases the calcium ion influx through excitable membrane; this is not affected by tetrodotoxin or sodium. Usually Maitotoxin is less important since it is less present in fish. Scaritoxin is similar to Ciguatoxin. Okadaic Acid is a lipid soluble toxin with a LD50 210 ug/kg ip in mice; it is a sodium ionophore. Palytoxin is a water soluble polyether which causes severe tonic contraction of all muscle groups; it also strong skin irritant and potent tumor activator.
The pharmacologic action of Ciguatoxin is due to its direct effects on excitable membranes. Its potent depolarizing action due to a selective increases in sodium permeability in the nerve cells and striated muscle can be counteracted by calcium ions and tetrodotoxin. The respiratory arrest induced by a lethal dose results mainly from depression of the central respiratory center. It causes biphasic cardiovascular response with hypotension and bradycardia (which can be antagonized with anticholinergics) followed by hypertension and tachycardia (which can be suppressed by adrenergic blockers). The response of smooth muscle to ciguatoxin is complex, depending upon the predominant autonomic innervation and postsynaptic receptor. It causes a potent release of endogenous norepinephrine from adrenergic nerve terminals and a potentiating effect on the post synaptic membrane.
Maitotoxin possesses a specific Ca2+ dependent action which causes a release of norepinephrine from rat pheochromocytoma cells. This action occurs in the absence of Na+ ions and in the presence of tetrodotoxin, precluding the participation of sodium channels; Maitotoxin appears to exert its effects on endogenous membrane calcium channels.
This information is courtesy of Lora E. Fleming, Director of the European Centre for Environment and Human Health and Chair of Oceans, Epidemiology and Human Health at the University of Exeter Medical School
Background
DSP is a gastrointestinal illness without neurologic manifestations reported worldwide (ILO 1984, Halstead 1988, Aune & Yndstad 1993). It is caused by the consumption of contaminated shellfish (Halstead 1988).
The first reported cases of DSP were in the Netherlands in the 1960s, followed by similar reports in the late 1970s from Japan (Aune & Yndstad 1993). Since then, more than 1300 cases have been reported from Japan, with the peak season from April to September. Other outbreaks have been reported in Europe and South America as well as the Far East. In Spain, over 5000 cases were reported in 1981; In France in 1984 and 1986, over 2000 cases were reported each year and over 300 cases were reported in Scandinavia in 1984 (Asomata et al, 1978, Yasumoto et al, 1980, Viviani 1992, Aune & Yndstad 1993). Mussels exported from Denmark to France caused DSP poisoning in over 400 people in 1990 (Hald et al, 1991). Finally in 1991 DSP was reported in over 100 people in Chile; in 1992, DSP was detected in toxic concentrations in shellfish beds in Uruguay (Lembeye et al, 1993, Mendez 1992, Aune & Yndstand 1993). Although DSP is reported worldwide, the most highly affected areas appear to be Europe and Japan (Aune & Yndstad 1993).
The causative organisms are the marine dinoflagellates Dinophysis, although there is an uneven distribution among species and location of toxin production. These dinoflagellates are widely distributed, but do not always form red tides. The associated toxins produced by the Dinophysis dinoflagellates are okadaic acid and its derivatives; there are at least 9 total toxins produced by these dinoflagellates. (Asomata et al, 1978, Yasumoto et al, 1980, ILO 1984, Lee 1989).
Clinical presentation
Diagnosis:
Management and Treatment:
Treatment is symptomatic and supportive with regards to short-term diarrhea and accompanying fluid and electrolyte losses. In general, hospitalization is not necessary; fluid and electrolytes can usually be replaced orally. Other diarrhetic illnesses associated with shellfish consumption, such as bacterial or viral contamination should be ruled out (Aune & Yndstad 1993).
As with many of the marine toxin induced diseases, the initial or index case(s) are often the tip of the iceberg. Therefore any suspected cases of DSP should be reported to the appropriate public health authorities for follow up to ascertain other cases and to prevent further spread. And every effort should be made to obtain contaminated materials and their source.
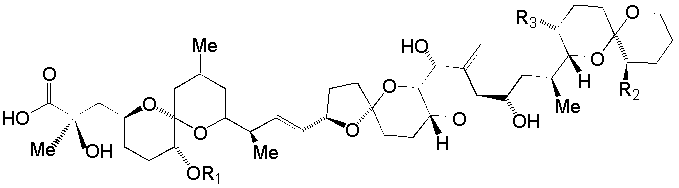{kind=link}
Molecular Mechanism of Action:
D. fortii at levels of 200 cell/litre in mussels and scallops becomes toxic for humans; the minimal amount of DSP toxins required to induce disease in humans was 12 MU.
Okadaic acid, dinophysiotoxin 1 and 3 are acidic, while there is another neutral group of toxins which are polyether lactones named pectonotoxins and yessotoxin. Diarrhea is caused in mice when the acidic component of okadaic acid is injected interperitoneally. Pectonotoxin 1 causes liver damage in mice under similar circumstances. Both the pectenotoxins and yessotoxin are lethal in mice with ip injection.
Okadaic acid is lipophilic. It is a potent inhibitor of protein phosphorylase phosphatase 1 and 2A in the cytosol of the mammalian cells that dephosphorylate serine and threonine. It probably causes diarrhea by stimulating the phosphorylation that controls sodium secretion by intestinal cells similar to Vibrio cholerae, although by a different mechanism. Okadaic Acid also acts through the variations of cellular concentration of the Ca2+ second messenger. It strongly increases the Ltype inward Ca2+ current in isolated guinea pig cardiac myocytes. Finally, okadaic acid functions not only as a tumor promotor (promoter of skin tumor in the mouse using DMBA as the initiator), but it is also capable of reversing cell transformation in some oncogenes.
References:
- AUNE T., and M. YNDSTAD: Diarrhetic shellfish poisoning. in: IR Falconer (Ed), Algal Toxins in Seafood and Drinking Water. London: Academic Press (1993) 87-104.
- Baden D, Fleming LE, Bean JA. Chapter: Marine Toxins. in: Handbook of Clinical Neurology: Intoxications of the Nervous System Part II. Natural Toxins and Drugs. FA deWolff (Ed). Amsterdam: Elsevier Press, 1995. pgs. 141-175.
- FUJIKI, H., M. SUGANUMA, H. SUGURI, S. YOSHIZAWA, K. TAKAGI, N. UDA, K. WAKAMATSU, K. YAMADA, M. MURATA, T. YASUMOTO and T. SUGIMURA: Diarrhetic shellfish toxins, DTX-1, is a potent tumor promoter on mouse skin. Japanese Journal of Cancer Research 79 (1988) 1089-1093.
- HAYSTEAD, T.A.J., A.T.R. SIM, D. CARLING, R.C. HONNOR, Y. TSUKITAN, P. COHEN and D.G. HARDY: Effects of the tumor promoter okadaic acid on intracellular protein phophorylation and metabolism. Nature 337 (1989) 78-81.
- LEE, J.S., Y. YANAGI, R. KENMA and T. YASUMOTO: Fluorometric determination of diarrhetic shellfish toxins by high performance liquid chromatography. Agricultural Biology and Chemistry 51 (1987) 877-881.
- LEE, J.S., M. MURATA and T. YASUMOTO: Analytical methods for determination of diarrhetic shellfish toxins. In: S Natori, K Hashimoto, Y Ueno (eds), Mycotoxins and Phycotoxins '88. Amsterdam, Elsevier (1989) 327-334.
- MURATA, M., M. SHIMATANI, H. SUGITANI, Y. OSHIMA and T. YASUMOTO: Isolation and structural elucidation of the causative toxin of the diarrhetic shellfish poisoning (1982) Bulletin of the Japanese Society of Scientific Fisheries 48: 549-552.
- YASUMOTO, T., Y. OSHIMA, W. SUGAWARA, Y. FUKUYO, H. OGURI, T. IGARASHI and N. FUJITA: Identification of Dinophysis fortii as the causative organism of diarrhetic shellfish poisoning. Bulletin of the Japanese Society of Scientific Fisheries 46 (1980) 1405-1411.
This information is courtesy of Lora E. Fleming, Director of the European Centre for Environment and Human Health and Chair of Oceans, Epidemiology and Human Health at the University of Exeter Medical School
Neurotoxic Shellfish Poisoning
In humans, two distinct clinical entities, depending on the route of exposure, are associated with exposure to the Florida Red Tide toxins. With the ingestion of contaminated shellfish, Neurotoxic Shellfish Poisoning (NSP) presents as a milder gastroenteritis with neurologic symptoms compared with paralytic shellfish poisoning (PSP). With the inhalation of the aerosolized Red Tide toxins, especially the brevetoxins, from the sea spray exposure associated with Florida Red Tide with and without accompanying fish kills, respiratory irritation and possibly other health effects in humans and other mammals occur (Baden 1995, Fleming 1998a&b, Fleming 1999, Bossart 1998).
Walker was the first to record NSP in 1880 on the West Coast of Florida. The associated red tides are often characterized by patches of discolored water, dead or dying fish and respiratory irritants in the air. Since then, NSP has been reported from the Gulf of Mexico, the East Coast of Florida, and the North Carolina coast. Recent prolonged red tides in the Gulf of Mexico have been associated with significant environmental, human health and economic impacts. Beaches in the Texas were closed, as well as shellfish beds from Florida to Texas. Significant die-offs of endangered manatees and double-rested cormorants, as well as reported human health effects, resulted secondary to the inhalation of the Red Tide toxins (Bossart 1998, Hopkins 1997, Kreuder 1998).
Organism & Toxins
The classic causative organism, Gymnodinium breve, is a dinoflagellate restricted to the Gulf of Mexico and the Caribbean, although similar species occur throughout the world. It is found especially during red tides in the late summer and autumn months almost every year off the West Coast of Florida with massive fish and bird kills. Fish kills associated with these red tides have been estimated up to 100 tons of fishes per day. Recently, these red tides are increasing in incidence, time length and geographic spread. Although the possibility of anthropogenic influences such as nutrient run-off is being investigated, it should be noted that these red tides in Florida occurred even prior to significant pollution from human population (Tester 1997, Tester 1991).
G. breve produces 2 types of lipid soluble toxins: hemolytic and neurotoxic. The neurotoxic toxins are known as brevetoxins. The major brevetoxin produced is PbTx-2; lesser amounts of PbTx--1, PbTx-3, and hemolytic components are produced. The massive fish kills are due to the neurotoxin exposure, with possible contribution of the hemolytic fraction. As with all the marine toxins, the brevetoxins are tasteless, odorless, and heat and acid stable. These toxins cannot be easily detected nor removed by food preparation procedures (Baden 1993).
The G. breve organism is relatively fragile. Therefore, particularly in wave action along beaches, the organism is easily broken open, releasing the toxins. During an active in-shore red tide, the aerosol of contaminated salt spray will contain the toxins and organism fragments both in the droplets and attached to salt particles; this can be carried in land depending on wind and other environmental conditions (Pierce 1990, Pierce 1989).
Molecular Mechanism of Action:
Fish, birds and mammals are all susceptible to the brevetoxins. The mouse LD50 is 0.20 mg/kg body weight (0.15-0.27) intraperitoneally. In human cases of NSP, the brevetoxin concentrations present in contaminated clams have been reported to be 30-18 ug (78-120 ug/mg).
The brevetoxins are lipid soluble polyethers with molecular weights around 900. These toxins are depolarizing substances that open voltage gated sodium (Na+) ion channels in cell walls, leading to uncontrolled Na+ influx into the cell (Baden 1983). This alters the membrane properties of excitable cell types in ways that enhance the inward flow of Na+ ions into the cell; this current can be blocked by external application of tetrodotoxin (Gallagher 1980, Baden 1983, Halstead 1988, Poli 1986, Viviani 1992, Trainer 1991).
It is believed that the respiratory problems associated with the inhalation of aerosolized Florida Red Tide toxins are due in part to the opening of sodium channels by the brevetoxin (Baden 1993, Asai 1982, Borison 1980, Franz 1989). In sheep, Abraham found that the bronchospasm can be blocked by atropine (verbal communication). In addition, there appears to be a role for mast cells; in sheep, the bronchospasm can be effectively blocked by cromolyn and chlorpheniramine (W Abraham PhD, verbal communcation). Watanabe et al (1988) noted that brevetoxin can combine with a separate site on the h gates of the sodium channel, causing the release of neurotransmitters from autonomic nerve endings. In particular, this can release acetylcholine, leading to smooth tracheal muscle contraction, as well as massive mast cell degranulation.
Since brevetoxins are also enzymatic inhibitors of the lysosomal proteinases known as cathepsins found in phagocytic cells such as the macrophages and lymphocytes, it is also possible that acute and chronic immunologic effects (including the release of inflammatory mediators that culminate in fatal toxic shock) may be associated with exposure to aerosolized Red Tide toxins, especially with chronic exposure and/or susceptible populations (Bossart 1998), although recent work by Baden et al cast doubt on the cathepsin mechanism (D Baden, verbal communication).
Clinical Presentation:
The two forms of Red Tide toxins-associated clinical entities first characterized in Florida are an acute gastroenteritis with neurologic symptoms following ingestion of contaminated shellfish (i.e. NSP), and an apparently reversible upper respiratory syndrome following inhalation of aerosols of the dinoflagellate toxins (i.e. aerosolized red tide toxins respiratory irritation) as well as possibly other human health effects (Baden 1995, Fleming 1999, Fleming 1998a, Fleming 1998b, Morris 1991).
Aerosolized red tide toxins respiratory irritation consists of conjunctival irritation, copious catarrhal exudates, rhinorrhea, nonproductive cough, and bronchoconstriction with the inhalation of the aerosol of Florida Red Tides, the toxins of G. breve. Some people also report other symptoms such as dizziness, tunnel vision and skin rashes. In the normal population, the irritation and bronchoconstriction are usually rapidly reversible by leaving the beach area or entering an air conditioned area (Steidinger 1984, Baden 1983). However, asthmatics are apparently particularly susceptible, a finding confirmed in recent investigations with an asthmatic sheep aerosolized Red Tide toxins respiratory irritation model (W Abraham, verbal communication). Furthermore, there are anecdotal reports of prolonged lung disease, especially in susceptible populations such as the elderly or those with chronic lung disease. Of note, the Red Tide toxins inhalation manatee die-off investigation in 1996 revealed severe catarrhal rhinitis, pulmonary hemorrhage and edema, and non-suppurative leptomeningitis, as well as possible chronic hemolytic anemia with multi-organ hemosiderosis and evidence of neurotoxicity in the dead manatees (Bossart 1998).
Exposure
Traditionally, aerosolized Red Tide toxins respiratory irritation is associated with significant Florida Red Tide blooms (including significant fish kills with dead fish on the beaches). Nevertheless, exposure to aerosolized Red Tide toxins can cause respiratory irritation in even non-asthmatics even without obvious fish kills or high dinoflagellate cell counts in the seawater within a few feet of the seashore (K. Steidinger, Florida Dept of Environmental Protection, verbal communication). The symptoms in non-asthmatic persons usually end rapidly within a few hundred feet of the seashore or upon entering significantly air-conditioned cars or homes. Also of interest, reportedly research has shown that the bevetoxins can be highly concentrated in the aerosol of sea spray generated by waves hitting the shore during a Red Tide (Pierce 1990, Pierce 1989). It is not known how far inshore this Red Tide toxins aerosol will travel, especially given strong off-shore winds during a Red Tide bloom. Although water sampling for both the dinoflagellates and the toxins has been performed for many years, Red Tide toxins air monitoring is presently experimental. Air monitoring could provide qualitative and quantitative time- and geographic-based data.
Diagnosis:
The diagnosis of Florida Red Tide toxins-associated clinical entities has been based on the clinical scenario of persons becoming ill with gastrointestinal and neurologic symptoms after eating shellfish or with acute respiratory symptoms similar to asthma after inhaling aerosols associated with exposure to Florida Red Tide toxins. There is a mouse bioassay with crude toxic residue extracted with ethyl ether and a mosquito fish bioassay. Recent promising research includes: an FPLC methodology for the identification of the G. breve toxins, as well as antibodies to brevetoxin and a possible cell based assay (Templeton 1988, Melinek 1994, Fairey 1997, Ishida 1996, Whitney 1997, Poli 1995).
Work with Florida manatees (apparently killed due the inhalation of the Red Tide toxins) has lead to the development of a qualitative immunocytochemical stain for the Florida Red Tide toxins found within the macrophages and lymphocytes in nasal mucosa, lung and other tissues (Bossart 1998). This technique has also been used in marine birds exposed to red tide toxins (Jessup 1998, Kreuder 1998). This biomarker can be used as both an indicator of exposure and effect. Based on recent research in a sheep animal model using a modified immunocytochemical technique on the bronchial lavage specimens of animals exposed to aerosolized red tide toxins, this biomarker holds promise as a diagnostic and prognostic tool. Initial work shows that the immunocytochemical staining of throat and nasal swab specimens reflect the bronchial lavage results, thus allowing for a more human-applicable biomarker.
Management and Treatment:
In the case of aerosolized Red Tide toxins respiratory irritation, the use of particle filter masks or retreat to air conditioned environment will anecdotally provide relief from the airborne irritation. In sheep exposed to aerosolized red tide toxins, the use of cromolyn or chlorpheniramine may treat, and if used prophylactically, even prevent the bronchoconstrictive response; this may have implications for asthmatics and other susceptible persons exposed to aerosolized Red Tide toxins (W. Abraham, verbal communication).
The Florida Department of Environmental Protection (DEP) since the mid 1970s has conducted a control program with the closure of shellfish beds when G. breve concentrations are greater than 5000 cells/liter, until 2 weeks by testing for toxin with mouse bioassay testing. This should prevent cases of ingestion NSP related to contaminated shellfish consumption in most of the Florida human population, but not the respiratory irritation associated with exposure to aerosolized Red Tide toxins. There is monitoring of these red tides with their characteristic discoloration and massive fish kills by the Florida DEP, as well as unsolicited reports of respiratory irritation to the Florida Dept of Health. Although other states such as Texas have done otherwise, in Florida where the Red Tides are almost a yearly occurrence, beaches are not closed to recreational or occupational activities, even during very active near-shore blooms.
In 1999, the Florida Dept of Health added NSP to their list of reportable diseases; however, aerosolized Red Tide toxins respiratory irritation is not a reportable illness. The Florida Poison Information Center at the University of Miami initiated a toll free 24 hour/day Marine Hotline (1-888-232-8635) in 1997 to increase the reporting of marine related illness, including the marine toxin associated diseases; any cases of reportable illnesses are passed on by the Poison Information Center to the Florida Dept of Health for official reporting purposes. The Poison Control Center specialists have received specialized training in the recognition and triage of the marine toxin related illnesses through CDC funding for Estuarine Associated Syndrome. Efforts are on-going to increase knowledge and reporting of these illnesses by healthcare providers and public health officials, including a Video Conference on the Human Health Effects of Marine Toxins in Florida in June 1999 with a video and educational materials by the NIEHS Center through funding from CDC, the Florida Dept of Health and AHEC.
Identified Research Areas
There is very little published literature or formal epidemiologic studies on the human health effects of the diseases, either ingestion NSP or inhalation aerosolized Red Tide toxins respiratory irritation. As a non-reportable disease, NSP is highly under--reported and under-diagnosed; for example, there are no existing statistics for the incidence of NSP or aerosolized Red Tide toxins respiratory irritation, even in endemic areas, nor on possible chronic health effects in humans. There are no established biomarkers for either of the Florida Red Tide toxins-associated conditions in humans, nor have there been any formal published studies of aerosolized Red Tide toxins respiratory irritation surveillance monitoring. There is very little information on appropriate treatment and prevention methodologies (Fleming 1995, Fleming 1998, Fleming 1999).
References:
- Asai, S., I.I. Krzanowski, W.H. Anderson, et al. 1982. Effects of the toxin of red tide, Ptychodiscus brevis, on canine tracheal smooth muscle: a possible new asthma triggering mechanism. J. Allergy Clin. Immunol. 69: 4 18-428.
- Baden, D.G. 1996. Analyses of biotoxins (red tide) in manatee tissues. Miami: Report #MR148, Marine and Freshwater Biomedical Sciences Cener, National Institute of Environmental Health Sciences, Rosenstiel School of Marine and Atmospheric Sciences.
- Baden, D.G., K.S. Reins, R.E. Gawley, G. Jeglitsch, and D.J. Adams. 1993. The a-ring lactone of brevetoxin PbTx-3 is required for sodium channel orphan receptor binding and activity. Natural Toxins.
- Baden, D.G. and T.J. Mende. 1982. Toxicity of tow toxins from the Florida red tide marine dinoflagellate, Gymnodinium breve. Toxicon 20: 457-461
- Baden, D.G. 1983. Marine food-borne dinoflagellate toxins. Intemational Review of Cytology 82: 99-150.
- Baden, D.G., L.E. Fleming, and J.A. Bean. 1995. Chapter: Marine Toxins. in: Handbook of Clinical Neurology: Intoxications of the Nervous System Part H. Natural Toxins and Drugs. FA deWolf (Ed). Amsterdam: Elsevier Press. pgs. 141-175.
- Borison, H.L., S. Ellis, and L.E. McCarthy. 1980. Central respiratory and circulatory effect of Gymnodinium brevetoxin in anaesthetized cats. British Journal of Pharmacology 70: 249-256.
- Bossart, G.D., D.G. Baden, R. Ewing, B. Roberts, and S. Wright. 1998. Brevetoxicosis in Manatees (Tnchechus manatus latirostris) from the 1996 epizootic: gross, histopthologic and immunocytochemical features. Tox. Path. 26(2): 276-282.
- Cohen, J. 1988. Statistical Power Analysis for the Behavioral Sciences. Hillsdale, NJ: Lawrence Frlbaum Associates.
- Fairey, E.R., J.S. Edmunds, and J.S. Ramsdell. 1997. A cell based assay for brevetoxins, saxitoxins and ciguatoxins using a stably expressed c-fos-luciferase reporter gene. Anal. Biochem. 251: 129-32.
- Fleming, L.E., J.A. Bean, and D.G. Baden. 1995. Epidemiology and Public Health. In: Manual on Harmful Marine Microalgae. Hallegraeff, G.M., D.M. Anderson, and A.D. Cembella, eds. Denmark: UNESCO.
- Fleming L.E. and D.G. Baden. 1998. Neurotoxic Shellfish Poisoning: Public Health and Human Health Effects. White Paper for the Proceedings of the Texas Conference on Neurotoxic Shellfish Poisoning, Proceedings of the Texas NSP Conference, Corpus Christi (Texas), April 27-34.
- Fleming, L.E. and J. Easom. 1998. Seafood Poisonings. Travel Medicine 2 (10):1-8.
- Fleming, L.E. and J. Stinn. 1999. Shellfish Poisonings. Travel Medicine 3:1-6.
- Franz, D.R. and R.D. LeClaire. 1989. Respiratory effects of brevetoxin and saxitoxin in awake guinea pigs. Toxicon. 27: 647-654.
- Gallagher, P. and P. Shinnick-Gallagher. 1980. Effect of G. brevetoxin in the rat phrenic nerve diaphragm preparation. British Journal of Pharmacology 69: 367-372.
- Halstead, B.W. 1988. Poisonous and venomous marine animals of the world. Princeton: Darwin Press.
- Hemmert, W.H. 1975. The public health implications of Gymnodiniurn brevered tides, a review of the literature and recent events. Proceedings of the First International Conference on Toxic Dinoflagellate Blooms. Boston: MIT, pgs. 489-497.
- Hopkins, R.S., S. Heber, and R. Hammond. 1997. Water related disease in Florida: continuing threats require vigilance. J. Florida Med. Ass. 84: 441-445.
- Ishida, H., N. Muramatsu, H. Nukay, T. Kosuge, and K. Tzuji. 1996. Study on neurotoxic shellfish poisoning involving the oyster, Crassostrea gigas, in New Zealand. Toxicon. 34: 1050-3.
- Jessup, D.A., J. Ames, G. Bossart, J. Hill, B. Gonzales, and A. DeVogelaere. 1998. Brevetoxin as a cause of summer mortality in common murres (Uria aalge) in California. Proc. Int. Assoc. Aquatic Animal Med. San Diego, CA.
- Kreuder, C., G.D. Bossart, and M. Elle. 1998. Clinicopathologic features of an epizootic in the double-crested cormorant (Phalacrocorax auritus) along the Florida Gulf coast. Proc Wildlife Dis Assoc. Madison, WI.
- Melinek, R., K.S. Rein, D.R. Schultz, and D.G. Baden. 1994 Brevetoxin PbTx-2 immunology: differential epitope recognition by antibodies from two goats. Toxicon. 32: 883-90.
- Morris, P., D.S. Campbell, T.J. Taylor and J.I. Freeman. 1991. Clinical and epidemiological features of neurotoxic shellfish poisoning in North Carolina. American Journal of Public Health 81: 471-3.
- Music, S.I., J.T. Howell, and L.C. Brumback. 1973. Red tide: its public health implications. Florida Med. J. 60(11): 27-29.
- Pierce, R.H., M.S. Henry, L.S. Proffitt and P.A. Hasbrouck. 1990 Red tide toxin (brevetoxin) enrichment in marine aerosol. Toxic Marine Phytoplankton. (E. Graneli, S. Sundstron, L. Elder and D.M. Anderson, eds.) pp. 397-402.
- Pierce, R., M. Henry, S. Boggess and A. Rule. 1989. Marine toxins in bubble-generated aerosol. In: The Climate and Health Implications of Bubble-Mediated Sea-Air Exchange (E. Monahan and P. van Patton, eds.), Connecticut Sea Grant Publications: 27-42.
- Poli, M., K.S. Rein, and D.G. Baden. 1995. Radioimmunoassay for PbTx2 type brevetoxins: epitope specificity of two anti-PbTx sera. J. AOAC International 78: 538-542.
- Poli, M., T.J. Mende, and D.G. Baden. 1986. Brevetoxins, unique activators of voltage-sensitve sodium channels bind to specific sites in rat brain synaptosomes. Molecular Pharmacology 30: 129-135.
- Steidinger, K.A. and D.G. Baden. 1984. Toxic marine dinoflagellates. In: D.L. Spector (ed), Dinoflagellates. New York: Academy Press, pp. 201-261.
- Templeton, C.B., M.A. Poli, and R.D. LeClaire. 1988. Antibody to prevent the effects of brevetoxin poisoning in conscious rats. Gov. Rep. Announce. Index 17.
- Tester, P. and K.A. Steidinger. 1997. Gymnodinium brevered tide blooms: initiation, transport and consequences of surface circulation. Limnol. Oceanogr. 45: 1039-1051.
- Tester, P.A., R.P. Stumpf, F.M. Vukovich, P.K. Fowler, and J.T. Turner. 1991. An expatriate red tide bloom: transport, distribution and persistence. Limnol. Oceanogr. 36: 1053-1061.
- Trainer, V.L., W.J. Thomsen, W.A. Catterall, and D.G. Baden. 1991. Photoaffinity labeling of the brevetoxin receptor on sodium channels in rat brain synaptosomes. Molecular Pharmacology 40: 988-994.
- Viviani, R. 1992. Eutrophication, marine biotoxins, human health. Science for the Total Environment -Supplement 631-62.
- Watanabe, T., R.F. Lockey and J.J. Krzanowski. 1988. Airway smooth muscle contraction induced by Ptychodiscus brevis (red tide) toxin as related to a trigger mechanism of bronchial asthma. Immuno. Allergy Pract. 10(5): 185-192.
- Whitney, P.L., J.A. Delgado and D.G. Baden. 1997. Complex behavior of marine animal tissue extracts in the competitive binding assay of brevetoxins with rat brain synaptosomes. Nat. Toxins 5: I 93-200.
This information is courtesy of Lora E. Fleming, Director of the European Centre for Environment and Human Health and Chair of Oceans, Epidemiology and Human Health at the University of Exeter Medical School
Background
PSP is a marine toxin disease with both gastrointestinal and neurologic symptoms reported worldwide. It is caused predominantly by the consumption of contaminated shellfish.
Gonyaulacoid dinoflagellates are the source of PSP marine toxins.These unicellular dinoflagellates develop algal blooms throughout the world for unknown reasons, although a variety of factors have been studied, including change in weather, upwellings, temperature, turbulence, salinity, and transparency. However, significant epidemics of PSP can occur in humans in the absence of a known red tide (Rodrigue et al. 1990). These dinoflagellates produce at least 12 toxins which are tetrahydropurines, and heat and acid stable. Saxitoxin was the first characterized and the best understood.
The major transvector for PSP are the bivalve molluscs (mussels, clams, oysters, with the Alaskan butterclam having the highest concentrations) (Sommer & Meyer 1937). PSP toxins are also found in certain crabs and snails which feed on coral reef seaweed. The transvectors accumulate the toxins via feeding in their digestive organs and soft tissues, apparently without harm to the transvectors.
Humans, birds and fish can all be affected by PSP toxins. Herbivorous zooplankton is the primary transvector which can in turn transmit the toxin to fish and possibly other marine creatures which consume zooplankton (Baden 1983). The usual route for humans is the consumption of raw or cooked contaminated shellfish. There has been only one case of human contamination through consumption of contaminated fish and bird kills in Indonesia. In this case, the whole fish was consumed including the viscera which could be contaminated with PSP from shellfish consumption by the fish prior to death (MacLean & White 1989, Viviani 1992).
Clinical Presentation:
Ingestion of molluscs contaminated with PSP results in the following clinical picture (Bower et al, 1981, Kao 1993). Five to 30 minutes from consumption, there is slight perioral tingling progressing to numbness which spreads to face and neck to moderate cases. In severe cases, these symptoms spread to the extremities with incoordination and respiratory difficulty. There are medullary disturbances in severe cases evidenced by difficulty swallowing, sense of throat constriction, speech incoherence or complete loss of speech, as well as brain stem dysfunction. Within 2-12 hours, in very severe cases, there is complete paralysis and death from respiratory failure in absence of ventilatory support. After 12 hours, regardless of severity, victims start to recover gradually and are without any residual symptoms within a few days (Bower et al, 1981, ILO 1984, Halstead 1988).
Other symptoms include headache, dizziness, nausea, vomiting, rapid pain, and anuria. There is no loss of consciousness and the reflexes are unaltered except maybe pupillary size and sight may be temporarily lost. As opposed to tetrodotoxin poisoning, there is rarely significant hypotension. Symptomatology is essentially identical for Pacific and Atlantic cases, although gastrointestinal symptoms may be more prominent in the Atlantic (ILO 1984, Halstead 1988).
The overall mortality (case fatality rate) was about 8.5% -9.5% in two large series (Meyer 1953, Ayres and Cullum 1978). However, the Guatemalan 1987 outbreak on Pacific coast had a case fatality rate of 14%, which was even higher in young children (50%). It is possible that children may be more sensitive to PSP toxins than adults (Rodrigue et al. 1990). In addition, the access to emergency medical services in acute cases is crucial to the prognosis.
The differential diagnosis of this clinical scenario of an acute gastrointestinal illness with recent shellfish ingestion would be bacterial or viral gastroenteritis. The neurologic manifestations are more consistent with poisoning by other marine toxins such as NSP and pufferfish poisoning, or even recent organophosphate pesticide poisoning.
Diagnosis:
The clinical scenario is the primary method of diagnosis initially. Recent shellfish ingestion, often but not always associated with known red tide, and an acute gastrointestinal illness with neurologic symptoms are part of the classic presentation. It is imperative to obtain samples of contaminated tissues and their source.
Each PSP epidemic is associated with different mixtures of the PSP toxins; this complicates the laboratory analysis of contaminated tissues. The mouse bioassay (time to death) of food extract is the recommended diagnostic method, (Sommer & Meyer 1937, Association of Official Analytical Chemists 1980) but it cannot distinguish between tetrodotoxin and other PSP toxins. The oral dose in humans for death is 1 to 4 mg (5,000 to 20,000 mouse units) depending upon the age and physical condition of the patient (see below). A mouse unit [MU] is defined as the minimum amount needed to cause the death of an 18 to 22 g white mouse in 15 minutes (Wiberg & Stephenson 1960, Shimizu 1984, Winter et al, 1990).
Radioimmunoassay and indirect enzyme-linked immunoabsorbent assay (ELISA) have been developed for saxitoxin but not all PSP toxins (Carlson et al, 1984). HPLC analysis method for all the PSP toxins has been developed with good correlation with mouse bioassay in terms of quantification (Sullivan et al, 1983, Halstead 1988).
Management and Treatment:
In general, supportive measures are the basis of treatment for PSP, especially ventilatory support in severe cases. In animals, artificial respiration is the most effective treatment. Without supportive treatment, upto 75% of severely affected persons die within 12 hours. Use of anticholinesterase agents are not recommended, and could actually be harmful (Murtha 1960, ILO 1984, Halstead 1988, Brown & Shepherd 1992, Kao 1993).
When the ingestion of contaminated food is recent, gut decontamination by the gastric lavage and administration of activated charcoal or dilute bicarbonate solution is recommended. Care must be taken concerning aspiration with the neurologically compromised patient. Anticurare drugs were ineffective, while DL amphetamine (benzedrine) was most effective in aiding the artificial respiration and decreasing the recovery period. Use of anticholinesterase agents are not recommended, and could actually be harmful (Murtha 1960, Bower et al, 1981, ILO 1984, Halstead 1988, Brown & Shepherd 1992, Kao 1993).
The lactic acidosis of unknown origin seen in experimental animals and possibly humans can be treated by assisted ventilation, fluid therapy and periodic monitoring of the blood pH. It is possibly that the fluid therapy will also assist in the renal excretion of toxin (Kao 1993).
Many endemic areas have traditionally used local treatments with variable success. In the Philippines, a drink of coconut and brown sugar is administered; demonstrations in mice show that these ingredients may have active detoxification substances (Viviani 1992).
As with many of the marine toxin induced diseases, the initial or index case(s) are often the tip of the iceberg. Therefore any cases of PSP should be reported to the appropriate public health authorities for follow up to ascertain other cases and to prevent further spread. And every effort should be made to obtain contaminated materials and their source.
Obviously the most effective form of PSP prevention is to eliminate human contact with contaminated shellfish and other transvectors. Surveillance and closures of commercial shellfish beds by monitoring the amount of PSP using the mouse assay are common practice throughout the world. For example, in the USA, PSP levels in edible shellfish greater than 800 ug PSP/kg by mouse assay means that commercial beds will be closed until they are monitored below this level; this action level is more than 10 times lower than the lowest level associated with human outbreaks [Anon 1965, ILO 1984]. Furthermore, there is active monitoring of algal blooms with fish and bird kills.
Ozonation can remove low levels of toxins from soft-shell clams but not if the clams have retained toxin for long periods of time; some industrial canning processes may lead to a decrease in PSP concentration (Halstead 1988, Viviani 1992). Biological controls such as using parasitic dinoflagellates to attack the red tide (for example Amoebophrya ceratii parasitizes a variety of dinoflagellates responsible for PSP) have been considered (Viviani 1992).
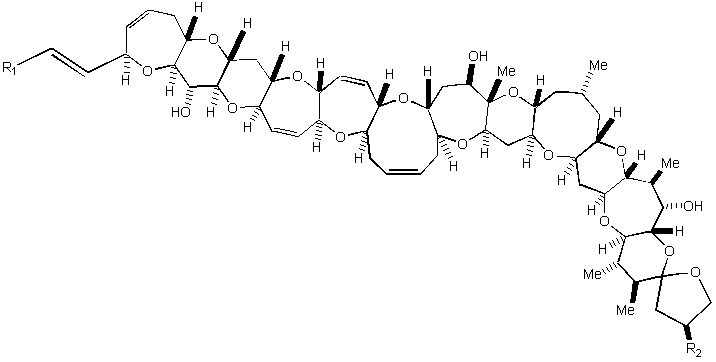{kind=link}
Molecular Mechanism of Action:
Saxitoxin is the most well known of the PSP associated toxins. It is a heat stable neurotoxin. In mice, the saxitoxin LD50 parentally is 3-10 ug/kg body weight and orally is 263 ug/kg body weight (death within minutes of respiratory failure). Humans are the most sensitive to saxitoxin; the oral dose in humans for death is 1 to 4 mg (5,000 to 20,000 mouse units) depending upon the ge and physical condition of the patient. It is rapidly absorbed through the gastro-intestinal tract and excreted in the urine.
Saxitoxin inhibits the temporary permeability of Na+ ions by binding tightly to a receptor site on the outside surface of the membrane very close to the external orifice of the sodium channel. In fact, neurophysiologic studies using saxitoxin as a probe helped to show that Na+ and K+ act independently with separate membrane channels. It is a blocking agent that reduces the number of conducting Na+ channels by occupying some site near the outer opening in a 1:1 high affinity specific receptor binding. This prevents sodium ions from passing through the membranes of nerve cells, thus interfering with the transmission of signals along the nerves. The resulting widespread blockade prevents impulse-generation in peripheral nerves and skeletal muscles. Saxitoxin has a direct effect on skeletal muscle by blocking the muscle action potential without depolarizing cells; it abolishes peripheral nerve conduction but with no curare-like action at the neuromuscular junction.
References:
- ASSOCIATION OF OFFICIAL ANALYTICAL CHEMISTS(AOAC). Paralytic Shellfish Poison Biological Method. In: Official Methods of Analysis. Washington, AOAC (1980) 289-299.
- AYRES, P.A. and M. CULLUM: Paralytic Shellfish Poisoning. London Ministry of Agriculture, Fisheries and Food(Fisheries Research Technical Report #4) (1978) 23.
- Baden D, Fleming LE, Bean JA. Chapter: Marine Toxins. in: Handbook of Clinical Neurology: Intoxications of the Nervous System Part II. Natural Toxins and Drugs. FA deWolff (Ed). Amsterdam: ElsevierPress, 1995. pgs. 141-175.
- CHENG, H.S., S.O. CHUA, J.S. HUNG and K.K. YIP: Creatinine kinase MB elevation in paralytic shellfish poisoning. Chest99 (1991) 1032-3.
- FRANZ, D.R. and R.D. LECLAIRE: Respiratory effects of brevetoxin and saxitoxin in awake guinea pigs. Toxicon27 (1989) 647-654.
- HALSTEAD, B.W. and E.J. SCHANTZ: Paralytic Shellfish Poisoning. Geneva, World Health Organization(1984) 60.
- KAO, C.Y.: Paralytic Shellfish Poisoning. In: IR Falconer (ed), Algal Toxins in Seafood and Drinking Water. London: Academic Press(1993)75-86.
- LONG, R.R, J.C. SARGENT and K. HAMMER: Paralytic Shellfish Poisoning: A case report and serial electrophysiologic observations. Neurology40 (1990) 1310-1313.
- MEE, L.D., M. ESPINOSA and G. DIAZ: Paralytic Shellfish Poisoning with a Gymnodinium catenatumRed tide on the Pacific Coast of Mexico. Marine Environmental Research (1986) 141-1136.
- RODRIGUE, D.C., R.A. ETZEL, S. HALL, E. DE PORRAS, O.H. VELASQUEZ, R.V. TAUXE, E.M. KILBOURNE and P.A. BLAKE: Lethal Paralytic Shellfish Poisoning in Guatemala. American Journal of Tropical Medicine and Hygiene42 (1990) 267-271.
- SHIMIZU, Y.: Paralytic shellfish poisons. Forthscritte Der Chemie Organischer Naturstoffe 45 (1984) 235-64.
- SULLIVAN, J.J., M.G. SIMON and W.T. IWAOKA: Comparison of HPLC and mouse bioassay methods for determining PSP toxins in shellfish. Journal of Food Science 48 (1983) 1312-1314.